Bats |
Seals |
Terrestrial Mammals
|
Whales and Dolphins
| |
From the 124 bat species covered by the GROMS, 3 are classified as "Endangered", 15 as "Vulnerable" and 17 as "Lower Risk/near threatened" by the International Red List 2000 (Hilton-Taylor 2000). Among these, the Mexican free-tailed bat (Tadarida brasiliensis) is the only species listed on CMS Appendix I (Figure A2.3). Some subspecies migrate between North and South America, and distances up to 1300 km have been observed. All European bat populations are listed on CMS Appendix II, and are covered by a CMS agreement on European bats (EUROBATS: Figure A2.2), regardless of their migratory status. Among them is the noctule bat Nyctalus noctula (Schreber 1774). Females migrate further than males, and North Eastern populations move up to 2000 km in south-western direction (Figure A2.4).
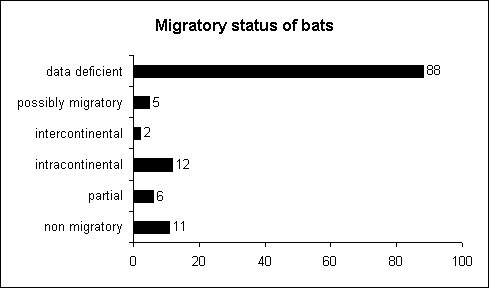
Baker (1978) has compiled maps and migration distances for 20 bat species in a compact format (l.c., pp. 566 ff.). They show a complex picture with considerable variation of migration distances between sexes and different populations (demes). As shown by Figure 4.4, there is still considerable uncertainty about the migratory status of bats. Most information about migration patterns is based on the
|
Fig. 4.4: Migratory status of 124 bat species listed by the GROMS. Non-migratory bats are included because they are listed on CMS Appendix II and covered by the EUROBATS agreement. Note the high number of species for which a reliable assessment was impossible. |
Abb. 4.4: Wanderverhalten der 124 von GROMS erfaßten Fledermausarten. Berücksichtigt wurden auch nicht wandernde Arten, da diese im CMS Anhang II enthalten sind und durch das EUROBATS Abkommen geschützt werden. Man beachte die hohe Zahl von Arten, für die eine zuverlässige Einteilung nicht möglich war. |
results of ringing studies, beginning with Eisentrautís extensive banding of Myotis myotis in minetunnels in Berlin from 1932 to 1950 (Eisentraut,1935). Roer (1995) has summarised the results of bat banding in Europe, and banding schemes are still carried out in Australia (Baker et al. 1997). However, banding has been criticised as being harmful for bats (Hutson 1999). Today, it is substituted and complemented by genetic fingerprinting (cf. Webb & Tidemann 1996; Wilkinson & Fleming 1996). Hopefully, such studies will help to improve our scanty knowledge, in particular about tropical migration patterns. The IUCN/SSC Chiroptera Specialist Group planned a symposium on migration for the 12th International Bat Research Conference (Malaysia 2001) with a workshop to identify potential and priorities for future research and conservation and the potential of the development of further international collaboration, especially in South America (Hutson 2001).
Migration patterns among New World bats are less well studied, but several species do exhibit transboundary migrations, mainly from the U.S. to Mexico, and therefore qualify for CMS listing (Table 4.2). These 4 species are all threatened because of "severe declines in area of occupancy" (criteria A1c, A2c), due to habitat destruction. The western long-nose bat (Leptonycteris curasoae), and in particular the subspecies L. c. yerbabuenae (lesser long-nosed bat), pollinates the saguaro, organ-pipe cacti and the agave, the plant from which tequila is made. It suffers from degradation of the Sonoran desert ecosystem, over-exploitation of Agave and modification or destruction of roost sites, which also affect its congener, the Mexican long-nosed bat (Leptonycteris nivalis). Both species are migratory. Wilkinson & Fleming (1996) suggest two migration routes for Leptonycteris curasoae, based on genetic analysis of unique haplotypes at distant sites, 1) along the Pacific coast ranging from at least Juxtlahuaca in Guerrero to south-western Arizona and (2) inland along the Sierra Madre possibly ranging from as far south as Chiapas to south-eastern Arizona. Leptonyceris nivalis (considered conspecific with L. curasoae). No such detailed studies are available for the other species listed in Table 4.2, but they all seem to migrate.
Several species of flying foxes (Pteropodidae: Megachiroptera) are known to migrate in response to flowering and fruiting of their fruit plants, but direct observation of movement patterns is difficult.
|
Tab. 4.2: Threatened migratory bat species, CMS status and migratory behaviour. Threat status is given according to Red List 2000 (Hilton-Taylor 2000). Note that only European populations are covered by the CMS through the EUROBATS agreement (Figure A2.2) |
Tab. 4.2: Bedrohte wandernde Fledermausarten, ihr Schutzstatus gemäß der Bonner Konvention und Wanderverhalten (Gefährdung nach Hilton-Taylor 2000). Man beachte, daß nur europäische Populationen durch die Bonner Konvention und das EUROBATS-Abkommen (Figure A2.2) geschützt sind. |
|||||
|
Family |
Scientific name |
Common name |
RL 2000 |
CMS |
Migration |
|
|
Mormoopidae |
Pteronotus macleayii |
MacLeayís moustached bat |
VU |
N.L. |
possibly |
|
|
Phyllostomidae |
Artibeus hirsutus |
Hairy Fruit-eating bat |
VU |
N.L. |
possibly |
|
|
Phyllostomidae |
Leptonycteris curasoae |
Western long-nosed bat |
VU |
N.L. |
intra- |
|
|
Phyllostomidae |
Leptonycteris |
Mexican long-nosed bat |
EN |
N.L. |
possibly |
|
|
Phyllostomidae |
Macrotus |
California leaf-nosed bat |
VU |
N.L. |
possibly |
|
|
Phyllostomidae |
Musonycteris |
Trumped-nosed bat |
VU |
N.L. |
data |
|
|
Rhinolophidae |
Rhinolophus |
Mediterranean horseshoe bat |
VU |
App. II (Pop) |
technical |
|
|
Rhinolophidae |
Rhinolophus |
Lesser horseshoe bat |
VU |
App. II (Pop) |
intra- |
|
|
Rhinolophidae |
Rhinolophus |
Mehelyís horseshoe bat |
VU |
App. II (Pop) |
technical |
|
|
Vespertilionidae |
Barbastella |
Barbastelle bat |
VU |
App. II (Pop) |
partial |
|
|
Vespertilionidae |
Myotis bechsteini |
Bechsteinís bat |
VU |
App. II (Pop) |
technical |
|
|
Vespertilionidae |
Myotis capaccinii |
Long-fingered bat |
VU |
App. II (Pop) |
data |
|
|
Vespertilionidae |
Myotis dasycneme |
Pond bat |
VU |
App. II (Pop) |
intra- |
|
|
Vespertilionidae |
Myotis |
Geoffroyís bat |
VU |
App. II (Pop) |
partial |
|
|
Vespertilionidae |
Myotis grisescens |
Grey myotis |
EN |
App. II (Pop) |
data |
|
|
Vespertilionidae |
Myotis sodalis |
Indiana bat |
EN |
N.L. |
intra- |
|
|
Vespertilionidae |
Plecotus |
Rafinesqueís big-eared bat |
VU |
N.L. |
data |
|
|
Vespertilionidae |
Plecotus |
Townsendís big-eared bat |
VU |
App. II (Pop) |
data |
|
Indirect evidence by an analysis of genetic subdivisions between populations indicates that Australian Pteropus spp. (e.g. Pteropus poliocephalus Grey headed flying-fox) should be considered migratory (Webb & Tidemann 1996), at least within large ranges in coastal north and eastern Australia. The African straw-coloured fruit bat (Eidolon helvum) occurs in large seasonal concentrations near Kampala, Uganda, but their migratory movements are not understood (Kingdon 1974).
Habitat degradation affects bat species worldwide. Destruction of roosting sites and cave colonies takes place even in remote areas, due to mining and cave tourism (cf. Johnson et al. 1997) Deliberate destruction of fruit-eating bat colonies is reported from Central American banana plantation, to prevent bats from leaving claw prints on fruit. Herbicides and pesticide application affect all species directly, and indirectly by destroying food resources for insectivorous bats. High organochlorine content in bat tissue has been reported worldwide (North America: Thies 1994, Thies et al. 1996, Thies & Thies 1997; Clark et al. 1987; Thailand: Kruthanut 1986; Spain: Fernandez et al. 1993), and in particular in industrialised countries due to roof timber treatment (Shore et al. 1989). For Germany, causes of threat have been carefully compiled by Boye et al. (1999). They list:
• Seals
This document is part of the publication "Riede, K. (2001): The Global Register of Migratory Species ≠ Database, GIS Maps and Threat Analysis. Münster (Landwirtschaftsverlag), 400 pp."
+ CD (see copyright), orders: