Mammalia |
Aves |
Reptilia
|
Pisces
| |
Birds are among the best-known organisms on earth. They are observed, counted and monitored to an extent unrivalled for any other animal group. Bird migration in particular has fascinated and puzzled humans since ancient times. Systematic ringing of birds was the method to find out where exactly migratory birds are wintering. Ringing data provided detailed distribution data, and impressive maps have been compiled in a multi-volume atlas about "Zug" (Zink & Bairlein 1995). In recent times, much progress has been made by satellite tracking, and some of the results are available on the Word Wide Web, as impressive visualisations of migratory paths of storks and other flagship species. In addition, new methods such as genetic fingerprinting and tracing of isotopes provided new insights about wintering ground of lesser-known species (Marra et al. 1998).
The physiological mechanisms underlying migration are still not fully understood. But the investigation of orientation, navigation and homing, as well as the underlying genetic programs, made enormous progress during the last decades. There is evidence for the use of a magnetic compass, solar and stellar orientation and landmarks. In addition, it is now becoming evident that several mechanisms interact, even within one species: birds probably switch from solar to magnetic orientation, even during one journey (Alerstam & Gudmundsson 1999). Amazing evolutionary adaptability of underlying genetic programs was demonstrated by Berthold (1998), who showed that the genetic component of migratory behaviour in the blackcap (Sylvia atricapilla) could be modified within few generations.
Not surprisingly, there is a considerable number of different definitions and classifications of bird migration. Among the most simple is distance, and the straightforward classification given by Boehning–Gaehse et al. (1998) was adopted by GROMS (see Table 2.1). However, further classifications are relevant, such as those based on migration behaviour (nocturnal migration, narrow front migration, wide front migration), or variation among populations. Salomonsen (1955) differentiated between populations from distinct breeding areas, but mixing in their wintering area, and those wintering in different areas (allohiemic populations). These finer subdivisions of migratory behaviour are not yet covered within the GROMS.
Besides this huge number of publications on basic mechanisms of "Zug", there is a vast literature on the ecology, phenology and biogeographic aspects. A bibliography on Neotropical migrants alone comprises 200 pages (Rappolle et al. 1998), and a search of bibliographic databases gives similar results. Luckily, there are several excellent summaries (Berthold 2000, Elphick 1995). It is estimated that each year 50 billion individuals travel, and that over half of the roughly 10,000 bird species migrates (Berthold 1998, p. 236). But these estimates depend on the definitions of migration used, and a number of 5,000 migratory bird species certainly includes small-scale migrations, such as along altitudinal gradients (see chapter 2).
Besides these fundamental biological studies, conservationists need quantitative data, which allow for estimations of mortality and reproductive rates. These data sets are still based on direct observations or ringing efforts by volunteers, organised in efficient monitoring networks. BirdLife International, together with its regional offices, has taken a lead for integrating these data for scientifically-based bird conservation, and compiles the International Red List Data presented above (BirdLife International 2000). Wetlands International is concentrating on "Waterbirds" and protection of their wetland habitats (see section 4.3.2.1). A net of monitoring stations strategically placed at important bottlenecks and staging areas (Figures A2.40, A2.75) provide data for large monitoring schemes, such as the "Mettnau-Reit-Illmitz-Program" of the Vogelwarte Radolfzell, Lake Constance, Germany (Berthold et al. 1986). Additional monitoring schemes operating in Europe have been listed by Marchant (1999), who discusses how these data on national level can be combined. Other large monitoring efforts, involving thousands of volunteers, are the North American "Christmas Counts" and "Breeding Bird Censuses" (Terborgh 1989). Additional valuable data sets are checklists for certain sites and areas, detailed atlases on a regional scale, and quantitative mapping by a raster approach (see section 4.4). Though there are still some "white spots" on the global map of monitoring sites, there is an overwhelming amount of data, which at least allows to envisage the "big picture" necessary for an effective conservation policy.
In spite of this wealth of information, coherent digital resources for the whole group are not readily available, and the higher systematics is still not settled. Most ornithologists do not accept the phylogeny by Sibley & Monroe (1991), which is based on DNA-analysis. The higher systematics outlined in Morony et al. (1975) is now widely accepted among conservationists, and used here. del Hoyo et al. (1992) compared both taxonomies, and also opted for the conservative scheme of Morony et al. (1975). These discussions could be ignored by conservationists, as phylogenies are hypotheses. However, a consistent use of higher systematics is decisive for CMS, because CMS Appendix II lists whole groups such as the songbird family Muscicapidae. Even scientific bird names in particular show major inconsistencies. According to general use by CMS and other conventions, GROMS follows Sibley & Monroe (1991, 1993) for species names. Some differences to other lists in use were noted, in particular when compared with standard sources such as "Handbook of the Birds of the World" (del Hoyo et al. 1992-2001). Therefore, the most important synonym is stored in a separate field of the species table, and listed explicitly within Annex I.
In the light of such inconsistencies, English vernacular names and "common group names" are sometimes more robust for retrieving digital information. Within the GROMS species report form, matching letters can be entered, and a search is performed within all common name fields, the scientific name, and the important synonym field. In addition, GROMS offers widely used categories such as "waterbirds" within the higher taxonomy tables of the database, which proved to be useful units for threat analysis and are used within this section (cf. Table 3.2 and Table 3.4).
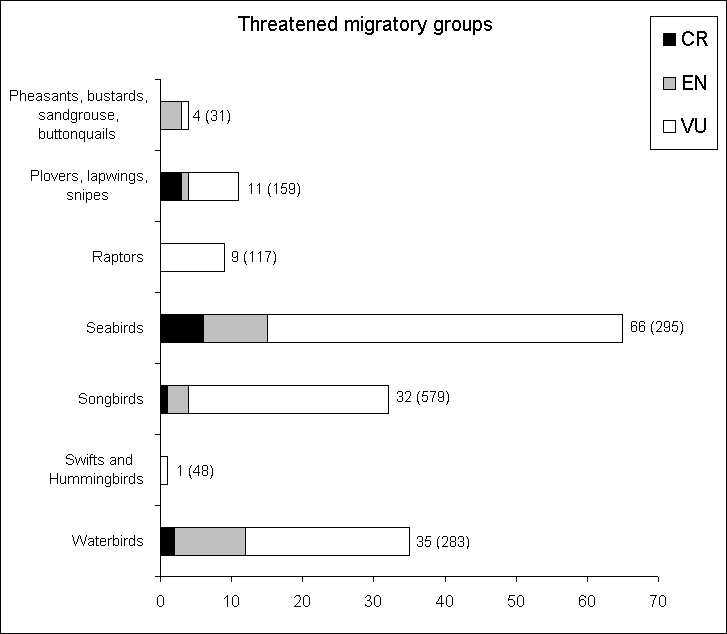
The summary statistics of threatened birds has already been discussed in section 4.2 (Figure 4.2). A more detailed analysis for major groups is presented in Figure 4.5, which shows the number of threatened migrants. Seabirds emerge as the most endangered groups, both with respect to absolute numbers and a proportion of approximately 20%, which is much higher than the average 12% for all birds. The following sections give a more detailed account of the conservation status of these major groups.
|
Fig. 4.5: Red list status according to IUCN Red List 2000 within major bird groups. Abbreviations see Figure 4.2. Numbers in brackets indicate total number of migrants for the group, as covered by the GROMS. Note the high number of threatened seabirds. |
Abb. 4.5: Gefährdungsstatus größerer Vogelgruppen gemäß der Internationalen Roten Liste 2000. Abkürzungen siehe Abb. 4.2. Zahlen in Klammern bezeichnen die Gesamtzahl wandernder Arten der jeweiligen Gruppe innerhalb des Weltregisters. Man beachte die hohe Zahl bedrohter Seevögel. |
Waterbirds and Plovers |
Seabirds |
Bustards
|
Raptors
|
Songbirds
|
Others
| |
This document is part of the publication "Riede, K. (2001): The Global Register of Migratory Species Database, GIS Maps and Threat Analysis. Münster (Landwirtschaftsverlag), 400 pp."
+ CD (see copyright), orders: